
 English
English


















„Kontroversa o neuroglii jest snad nejzajímavější ze všech, která se vedou v histologii nervstva.“ Tento výrok z r. 1899 patří českému biologovi a lékaři Vladislavu Růžičkovi (1870–1934) – viz AB 03/2009 – a je platný dodnes. Podle obecného povědomí se centrální nervový systém (CNS), jehož základním úkolem je získávání, zpracování a ukládání informací a odpověď na vnější podněty, skládá převážně z neuronů, které všechny tyto funkce zajišťují, ale skutečnost je poněkud jiná. Cílem stručného přehledu je napravit některé mylné představy o gliových buňkách a přiblížit čtenářům některé poznatky, které jejich funkci představí v úplně jiném, a možná překvapivém světle. Mnohé cenné údaje o gliových buňkách získali badatelé z Ústavu experimentální medicíny AV ČR.
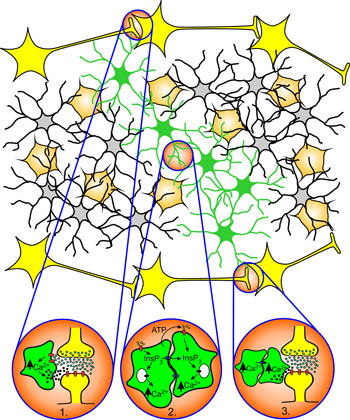
Zdroj: Archiv autora
Přenos informace v centrálním nervovém systému prostřednictvím propojených astrocytů:
1. Modulace synaptické aktivity neuropřenašečem uvolněným z astrocytů do štěrbiny aktivované synapse.
2. Mechanizmus pohybu „vápníkové“ vlny v síti z propojených astrocytů.
3. Modulace nebo aktivace vzdálené synapse neuropřenašečem uvolněným z astrocytů do synaptické štěrbiny.
Již v 18. století si švýcarský anatom a fyziolog Albrecht von Haller (1708–1777) všiml, že kromě ústrojí odpovědných za podráždění a čití, tj. neurony a nervová vlákna, je v mozku a míše přítomen další druh tkáně. Později tuto tkáň německý fyziolog a anatom Johannes Müller (1801–1858) pojmenoval jako pojivovou. Základní koncept nervové pojivové tkáně poprvé formuloval Purkyňův asistent Gabriel Valentin (1810–1883), jenž pozoroval nervová vlákna a nervové buňky obklopené další strukturou, a podle kterého byly tyto tři součásti specifickými útvary nervové soustavy tvořícími jeden celek. Patologickými změnami nervové pojivové tkáně se zabýval i český patolog a anatom Karel Rokytanský (1804–1878). V roce 1854 formuloval koncepci, podle níž byla nervová pojivová tkáň zásobárnou a kostrou, případně vazebnou hmotou pro specifické elementy nervové tkáně a rovněž základem a výchozí oblastí pro její regeneraci. Za „otce“ neurogliové koncepce nervové pojivové tkáně je však všeobecně považován německý anatom a patolog Rudolf Virchow (1821–1902); pojivová tkáň pozorovaná v mozku, v míše a v některých smyslových nervech byla podle něj jakýmsi druhem lepidla (pojmenoval ji jako „neuroglie“), ve kterém se nacházely nervové prvky. Určitým mezníkem ve výzkumu neuroglie se stala práce německého neuroanatoma Otto Deiterse (1834–1863). Ten jako první popsal v pojivové hmotě CNS buněčné elementy, které se odlišovaly od nervových buněk. V dalším období se počet badatelů studující vlastnosti gliových buněk zvýšil několikanásobně. Mezi nimi byl italský lékař a histolog Camillo Golgi (1843–1926), který v pojivové tkáni popsal okrouhlé, oválné a hvězdicovité buňky, mnohé z nich s výběžky. Výsledky C. Golgiho a jeho studentů, kteří v průběhu následného období identifikovali velké množství gliových buněk v bílé a šedé hmotě mozku a míchy, tvořily základ pro neurohistologický výzkum neuroglie konce 19. století. Německý neurohistolog a fyziolog Franz Boll (1849–1879) upozornil na rozmanitosti forem gliových buněk (nazýval je Deitersovými buňkami) a jako první se pokusil je rozdělit do skupin. Zhruba ve stejné době maďarský histolog a anatom Michael von Lenhossek (1863–1937) zavedl pojem „astrocyt“.
O další klasifikaci gliových buněk se zasloužili britský patolog William Andriezen (1870–1906) a švédský anatom Magnus Gustaf Retzius (1842–1919). Soustavnému studiu neuroglie se v té době v Praze věnovali český patolog Arnold Spina (1850–1918) a jeho student a budoucí český lékař Josef Vejnar (1867–1934), kteří popsali pohyb obarvených neurogliových buněk v šedé hmotě mozku žáby. Ten se mírným elektrickým prou-dem dal urychlit, avšak silným zastavit. Vejnar tyto změny popsal následovně: „Někdy pozorujeme, jak změny gliové přes zrnka se ubírají, což připomíná maně stín mračen přes dědiny přebíhající, nebo pohyb vlnícího se obilí kolem předmětu z něho vyčnívajícího.“ Pravděpodobně šlo o objemové změny neurogliových buněk. Další pokrok ve výzkumu astrocytů uskutečnil španělský histolog Santiago Ramón y Cajal (1852–1934), který prokázal, že v šedé hmotě se vyskytují především protoplazmatické astrocyty, zatímco vláknité (fibrilární) astrocyty jsou většinou v bílé hmotě. Ramón y Cajalův popis astrocytů a jejich výběžků a jejich vztahu k cévám a k neuronům zdokonalil názory na astrogliové buňky do podoby, jaká platí i v dnešní době. Kromě neuronů a astrocytů Ramón y Cajal studoval i další typ gliových buněk, který pojmenoval jako „třetí element“. Ten podrobně prozkoumal jeho student, argentinský histopatolog Pio del Rio Hortega (1882–1945) a popsal v něm dva druhy buněk, oligodendrocyty a mikroglie.
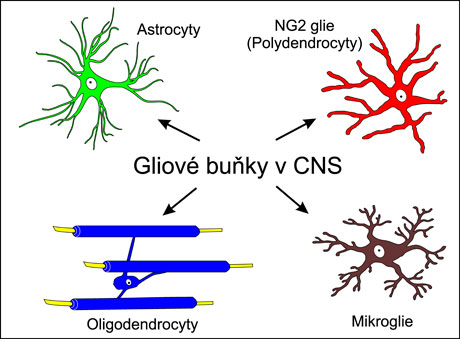
Uvedené výsledky tak tvoří jádro našich poznatků o buněčné organizaci neuroglie a jsou nedílnou součástí učebnic. Lze shrnout, že v CNS jsou přítomné čtyři typy gliových buněk. Tři z nich jsou ektodermového původu, tj. astrocyty, oligodendrocyty a nedávno charakterizované NG2-glie, zatímco čtvrtý typ, mikroglie, je mesodermového původu. Intenzivní výzkum prokázal, že gliové buňky se účastní fyziologických a patologických dějů a rozmach nových výzkumných metod, jako např. elektronová mikroskopie, elektrofyziologie (především metoda terčíkového zámku) a molekulární biologie, pomohl určit, že se gliové buňky mohou aktivně zapojit i do přenosu signálů v mozku a v míše.
Kolik je gliových buněk v CNS?
Na zdánlivě jednoduchou otázku není až tak snadné odpovědět. Důvodem je nejen přítomnost čtyř základních typů gliových buněk, ale i rozmanitost metod, které různí autoři použili pro určení jejich absolutního počtu v nervové tkáni. Zajímavým údajem však nejsou absolutní počty jednotlivých druhů gliových buněk, ale poměr mezi nimi a neurony. Všeobecně se považuje za prokázané, že u potkanů a myší je tento poměr 0,4 a u člověka 1,6. Nejvyšší je však u plejtváka malého – až 7,5. Pozoruhodné je, že zatímco morfologie neuronů u potkana a u člověka je podobná, lidské astrocyty jsou ve srovnání s potkany mnohem více rozvětvené, mají větší počet výběžků, jejich celkový objem je až 25krát větší a jejich výběžky směřují k 20krát většímu počtu spojení mezi neurony – synapsí. Zajímavé výsledky přineslo porovnání poměru glie/neurony u primátů a u člověka ve II. a III. vrstvě 9. prefrontální oblasti levé hemisféry, která odpovídá za některé vyšší kognitivní funkce, jako např. krátkodobá paměť, odhalování chyb, induktivní uvažování, empatie, zpracování emočních podnětů. Nej-menší poměr byl zjištěn u tamarína (0,46), u šimpanze byl již 1,20, a u člověka 1,65. V mozku člověka byly zjištěny i regionální rozdíly počtů neuronů a gliových buněk v mozku: v mozkové kůře člověka, která tvoří zhruba 82 % hmotnosti celého mozku, je přibližně čtyřikrát více gliových buněk než neuronů, zatímco v mozečku je gliových buněk přibližně čtyřikrát méně než neuronů.

Zdroj: Archiv autora
Vlastnosti astrocytů
Astrocyty jsou nejrozšířenějším typem gliových buněk v CNS. Většinou mají hvězdicovitý tvar, avšak podle novějších údajů je jejich morfologie mnohem rozmanitější. Dělí se na několik skupin, největší z nich tvoří protoplazmatické (plazmatické) astrocyty v šedé hmotě mozku a vláknité astrocyty v bílé hmotě. Další velkou skupinou je radiální glie, kterou tvoří bipolární buňky se dvěma dlouhými výběžky. Vyskytují se v největším počtu v počátečním období vývoje mozku, avšak v některých oblastech mozku přetrvávají až do dospělosti, jako např. Müllerovy buňky v sítnici oka a Bergmannovy buňky v mozečku. Kromě těchto hlavních skupin ještě existuje několik menších skupin specializovaných astrogliových buněk. Společnými hlavními vlastnostmi všech astrocytů je, že nevytvářejí akční potenciály, přestože na jejich membránách jsou přítomné ionotropní receptory pro neuropřenašeče (například glutamát, GABA, glycin), iontové kanály (K+, Na+, Ca2+, Cl-), metabotropní receptory (glutamát, GABA, ATP, adenozin) a specializované transportní mechanismy (např. pro GABA, glutamát a řadu iontů). Navíc, astrocyty vykazují některé morfologické zvláštnosti: prostřednictvím nízkoodporových spojů typu gap-junction jsou mnohdy propojeny do rozsáhlých sítí, tzv. funkčních syncytií.
Funkce astrocytů je velmi rozmanitá. V průběhu vývoje nervové tkáně napomáhají migraci neuronů a podílejí se na uspořádání různých vrstev šedé hmoty. Ve zralé tkáni poskytují strukturální oporu nervové tkáni. Astrocyty, jako jediné buňky v CNS, obsahují glykogen, který je zásobárnou energie pro správnou činnost neuronů. Perivaskulární astrocyty, které svými patkovitými výběžky zcela obalují cévy, se podílejí na tvorbě mozkomíšní bariéry a mohou dokonce regulovat průtok v cévách. Ependymocyty obklopují vnitřní komory mozku, jsou zapojeny do tvorby a sekrece mozkomíšního moku a svými kmitajícími řasinkami pomáhají jeho cirkulaci. Podílejí se na tvorbě hematoencefalické bariéry a předpokládá se, že mohou působit jako neurální kmenové buňky. Přítomnost iontových kanálů a specializovaných transportních mechanismů na membránách astrocytů jim umožňuje udržovat homeostázu mikro-prostředí neuronů, která se dramaticky mění v průběhu neuronální aktivity. Astrocyty mohou překvapivě uvolňovat do extracelulárního prostoru neuropřenašeče a neuromodulátory, a pokud tento děj probíhá v těsné blízkosti synapsí, může docházet k ovlivnění neuronálního přenosu. Ostatně, výběžky některých typů astrocytů synapse zcela obalují a jsou pravděpodobně schopny modulovat synaptickou aktivitu. Rovněž se prokázalo, že v syncytiích tvořených propojenými astrocyty se mohou šířit tzv. vápníkové vlny charakterizované krátkodobým nárůstem Ca2+ uvnitř buněk. Jde o velmi zajímavý jev, který pravděpodobně umožňuje synchronizovat aktivitu neuronů v blízkosti syncytia. Astrocyty se také mohou podílet na tvorbě nových synapsí a tím i ovlivňovat vyšší mozkové funkce, jako je spánek, paměť a učení.
Vlastnosti oligodendrocytů
Oligodendrocyty zahrnují dvě základní skupiny buněk. Jednak to jsou myelinizující oligodendrocyty, převážně v bílé hmotě, které v CNS vytvářejí na axonech neuronů izolační vrstvy z myelinu, přerušované tzv. Ranvierovými zářezy. Myelin je obsažen v útvarech připomínajících tenké destičky na konci krátkých výběžků, které se několikrát obtáčí okolo určitého úseku axonu. Tato izolace umožňuje na axonech tzv. saltatorní (skokové), a tudíž mnohem rychlejší šíření akčního potenciálu. Jeden oligodendrocyt může v CNS ve svém okolí obalit myelinovými pochvami až 30 axonů. Další skupinou jsou perineuronální (satelitní) oligodendrocyty, které se nacházejí převážně v šedé hmotě. Přisedají k tělům neuronů natolik těsně, že je možné na neuronech pozorovat prohlubeň. Jejich funkce je stále neznámá. Předpokládá se, že se mohou podílet na metabolismu neuronů a chránit je před apoptózou. Oligodendrocyty vykazují přítomnost celé řady ionotropních a metabotropních receptorů a iontových kanálů a předpokládá se, že jejich fyziologické funkce jsou shodné s funkcemi astrocytů.
Vlastnosti NG2-glie
NG-2 gliové buňky jsou někdy zařazovány mezi oligodendrocyty, neboť mají stejný původ, avšak na základě svých zvláštních vlastností jsou nyní vyčleněny do zvláštní skupiny. Byly charakterizovány teprve v 80. letech minulého století díky protilátkám proti chondroitinsulfát proteoglykanu NG2 (odtud pramení jejich název) a někdy bývají nazývány jako synantocyty nebo polydendrocyty. Jde převážně o hvězdicovité buňky, které se nacházejí v bílé i v šedé hmotě, nevytvářejí myelin a ve zralé tkáni se z nich vyvíjejí oligodendrocyty. Stejně jako oligodendrocyty vykazují přítomnost celé řady receptorů a iontových kanálů. Jejich zvláštností je, že vytvářejí funkční synapse s neurony a odpovídají na uvolnění neuropřenašečů do synapsí. Jde o stále málo probádané gliové elementy v CNS.
Vlastnosti mikroglie
Mikroglie jsou jediným typem gliových buněk mezodermového původu. Jsou to vlastně makrofágy, které do CNS pronikají ještě před uzavřením hematoencefalické bariéry, kdy se díky svému amébovitému pohybu podél cév a myelinizovaných vláken dostávají do všech oblastí nervové tkáně. Předpokládá se, že každá buňka zaujímá předem danou oblast. V klidu mají rozvětvené výběžky, zatímco při patologických stavech se opět mění na amébovité buňky s velkou schopností migrovat do místa poškození nebo infekce, proliferovat (množit) a fagocytovat rozpadlé zbytky tkáně nebo cizorodé částice. Mikroglie vykazují celou řadu iontových kanálů a receptorů pro neuropřenašeče, jejichž přítomnost závisí na funkčním stavu mikroglie. Zvláště důležitá je přítomnost receptorů pro cytokiny a chemokiny, které modulují pohyblivost a chemotaxi mikroglií. V poslední době se překvapivě prokázalo, že mikroglie se rovněž mohou uplatnit v průběhu vzniku nových synapsí během raného vývoje jedince. Rovněž tak mohou odstranit již nefungující nebo špatně fungující synapse.
Zdroj: Archiv autora
Úloha gliových buněk v průběhu patologických stavů
Významný vztah mezi nervovou pojivovou tkání (neuroglií) a patologickými stavy rozpoznal již Karel Rokytanský v roce 1854. Úloha gliových buněk v průběhu akutních a chronických patologických stavů se studuje i v současné době. Jsou to například mozkové mrtvice, anoxie, ischemie, degenerativní onemocnění CNS, jako Alzheimerova, Parkinsonova, Huntingtonova choroba a amyotrofická laterální skleróza, a nádory (gliomy). Vyjmenovat všechny patologické stavy spojené s gliovými buňkami by bylo nad rámec tohoto přehledu, proto jen několik příkladů.
Akutní poškození mozkové tkáně, například mechanické, doprovázejí výrazné změny ve složení extracelulárního prostoru, buněčná smrt a funkční i morfologické změny astrocytů, tzv. astroglióza. Aktivované astrocyty jsou schopny proliferovat v místě, kde je zasažena nervová tkáň, a jsou tak příčinou vzniku gliové jizvy, bariéry, která brání difuzi iontů, neuropřenašečů, trofických faktorů a dalších metabolitů z poškozené do zdravé nervové tkáně. Gliová jizva však bohužel zároveň brání regeneraci nervových vláken a představuje závažný problém při terapii např. míšních poranění.
Alzheimerovu neurodegenerativní chorobu vyvolává hromadění depozitů proteinu β-amyloidu v šedé hmotě a hromadění τ-proteinu uvnitř neuronů. Způsobuje rychlý úbytek neuronů v CNS s následným poškozením paměťových a kognitivních funkcí jedince a vznikem senilní demence. Bylo prokázáno, že neurodegenerace je doprovázena nejprve aktivací mikroglie a astrogliózou a poté úbytkem astrocytů, které jsou jinak schopny svými výběžky β-amyloidová depozita obalit a pohltit. Amyotrofickou laterální sklerózu doprovází degenerace motoneuronů v mozkové kůře, mozkovém kmeni a míše, která způsobuje progresivní paralýzu a atrofii svalů. Předpokládá se, že tato choroba je doprovázena astrogliózou a aktivací mikroglie. Je způsobena degenerací astrocytů a jejich omezenou schopností vychytávat glutamát z extracelulárního prostoru; následkem je buněčná smrt neuronů. Parkinsonovu nemoc charakterizuje progresivní degenerace dopaminergních neuronů v mozkové struktuře nazvané substantia nigra. Je doprovázena astrogliózou a aktivací mikroglie, o které se předpokládá, že by mohla mít významnou úlohu v průběhu degenerace neuronů. Prokázalo se totiž, že inhibitory aktivace mikroglie snížení počtu dopaminergních neuronů omezily. Huntingtonova choroba je neurodegenerativní onemocnění způsobené úbytkem neuronů v mozkové kůře a ve striatu. Patologická přítomnost astrocytů je spojena se sníženou expresí glutamátových transportérů, kdy následně extrémně stoupá koncentrace glutamátu a jeho homeostáza je narušena.
Nádory vznikající z gliových buněk jsou známy jako maligní gliomy. Jejich patologický potenciál spočívá v tom, že k šíření potřebují prostor, a tak ve svém okolí aktivně „zabíjejí“ neurony. Jedním z mechanismů je nadměrné uvolňování glutamátu na takovou hladinu, která již má cytotoxický účinek. Navíc, maligní buňky mají schopnost měnit svůj tvar a migrovat do zdravých oblastí nervové tkáně. Mohou rovněž vyloučením určitých faktorů ovlivnit mikroglie natolik, že je mikroglie již nepohlcují.
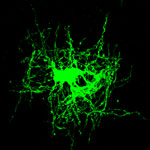
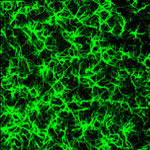
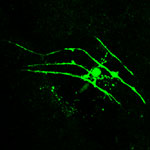
Zdroj: Archiv autora
Astrocyty a oligodendrocyt v míše potkana.
Obr. 1: fluorescenčně označený astrocyt.
Obr. 2: imunohisto-chemicky označené astrocyty v zadních rozích míchy.
Obr. 3: fluorescenčně označený oligodendrocyt.
Podle současných znalostí nejsou gliové buňky pouze podpůrnými elementy nervové tkáně, jak jsou ještě mnohdy vnímány. Na jednu stranu jsou strukturální oporou neuronů, podílejí se na tvorbě hematoencefalické bariéry a vytvářejí gliové sítě a myelinovou izolaci na axonech. Na straně druhé se na jejich membránách, podobně jako u neuronů, nachází množství napěťově závislých kanálů, chemicky aktivovaných iontových kanálů, metabotropních receptorů a transportních mechanismů. Gliové buňky mohou navíc uvolňovat některé neuropřenašeče a neuromodulátory do extracelulárního prostoru nebo do synaptických štěrbin a přispívat tak nejen k modulaci synaptického přenosu, ale i k plastickým změnám na neuronech. Neuroglie rovněž přispívá k zajištění homeostázy energetického metabolismu a k iontové a objemové homeostáze. V průběhu patologických změn gliové buňky hypertrofují, proliferují, vytvářejí gliální jizvy a podílejí se na případné remyelinizaci nebo na celkové regeneraci nervové tkáně.
Čím více toho o gliových buňkách víme, tím více se ukazuje, jak je struktura nervové tkáně složitá a jak je úloha gliových buněk v činnosti mozku a míchy nezastupitelná. Nejdůležitějším faktorem však zůstává, že gliové buňky a neurony tvoří jeden, vzájemně neoddělitelný funkční celek. Proto musíme na nervovou tkáň nahlížet jako na složitý organismus, který se skládá z různých typů buněk, a pouze jejich vzájemná spolupráce umožňuje normální fungování mozku a míchy a dokonce zajišťuje takové funkce, jako je naše vědomí.
ALEXANDR CHVÁTAL,
Ústav experimentální medicíny AV ČR, v. v. i.